
Notes on Mutation, Evolution and Selective Breeding
Just some further information that may be of interest to the more nerdy of my readers.
Already know what you're looking for? Quick Links |
More Stuff on the Workings of Genes
Genes code for proteins. Proteins make the world go round. Not quite literally, but certainly we wouldn't be here to notice the world going round if it wasn't for proteins. They are involved in every process within our cells and consequently enable life itself.
A protein consists of a chain of amino acids. There are 20 different types of amino acids, which makes for a lot of possibilities in the chains, but not every combination will produce a working protein. A gene is a set of instructions telling the cell exactly which amino acids should be present and in which order. The cell reads these instructions and assembles the correct amino acids in processes known as transcription and translation.
The exact structure of these instructions is a series of nucleotides, also known as bases, which are used as indicators (i.e. they tell the cell something). These bases are A (adenine), G (guanine), T (thymine) and C (cytosine). A sequence of three bases (a "codon") indicates a particular amino acid. There are also codons that indicate the end of the information for that gene (stop codons). In most DNA sequences there are actually long sections of redundant codons, which don't code for anything and are simply ignored during transcription. These sections are left over from previous points in the evolution of the species, and can actually be really useful clues for helping to work out genetic links between a species and its ancestors.
X and Y - Sex Chromosomes
Genes reside on structures of DNA called chromosomes, of which dogs have 39 pairs in their cells (humans have 23). The X and Y chromosomes determine gender, and contain a small number of genes. In mammals, females have two X chromosomes (XX) and males have one X and one Y (XY). The Y is the
male chromosome, and any mammal with just one Y will be male. YY is not a possible combination, as that could only be made from XY x XY - i.e. a breeding between two males!
The ratio of males to females is always roughly 1:1 (50% male, 50% female), as we can tell by drawing up a quick punnet square:

The Y chromosome actually contains far fewer working genes than the X chromosome. It can afford to have fewer - a male mammal will always have an X chromosome as well as a Y, so if a gene is on the X but not on the Y, it can just make use of the copy on the X. This means that there are some genes which are sex-linked, because they only occur on one of the sex chromosomes.
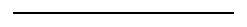

There are no X-linked colour genes in dogs, so the implications of this can be more easily seen in tortoiseshell (calico) cats. Tortoiseshell is a mixture of phaeomelanin (red tabby, in fact - cats don't come in the plain
red coats we see in dogs) and eumelanin, patched in a random pattern. The gene causing a cat to be red or black has two forms - O causes "orange" (red tabby, essentially works in a similar way to recessive red in dogs) and o (eumelanin, usually black). This gene is only carried on the X chromosome, so each cat will have either O or o on each of its X chromosomes. As male
cats only have one X, they only have one copy of the O allele. If that copy is is O then they will be solid red (or rather, red tabby), and if the copy is o then they will be solid black (or other eumelanin colour). But of course females have two Xs, so they have two copies of the O gene. If they are heterozygous (one copy of O and one copy of o, rather than two copies of one of those), they will be tortoiseshell.
The random patching is caused by a process where the female cat "switches off" one of her X chromosomes (chosen at random) in each of her cells
at some point during her early development. At this point she is still just a small embryo or foetus consisting of not very many cells (remember that when fertilisation of an egg cell occurs it immediately begins to multiply, so all cells making up the animal are descendants of that one cell). As she develops further the cells continue to multiply, and those that are descended from the cells where the X carrying o has been turned off will produce red pigment (phaeomelanin), because they're using the O allele on the active chromosome to determine the pigment protein they produce. And similarly, if the X carrying the O allele is turned off, the cells will produce black
pigment (eumelanin). The group of cells descended from each original cell will make up the patch of black or red. The size of the patches depends on how many
cells are descended from the original one where the switch-off took place. Hence, the later in development the switch-off occurs in a particular cell,
the smaller the patch associated with that cell will be. Which cells switch off which chromosome is entirely random and not determined by genetics, so even twin tortoiseshell kittens would have different patches. This has caused some surprise and confusion when tortoiseshell cats have been cloned in laboratories and the clone has ended up looking completely different to the original cat!
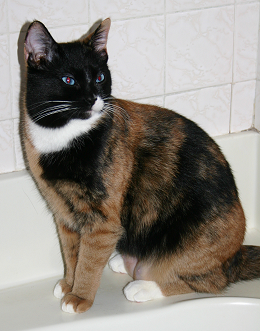


Believe it or not, all three of these tortoiseshell cats are from the same litter. Photos submitted by Chandra Mincher.
The system involving switching genes on and off is known as "epigenetics", and the process of switching off one of the X chromosomes is called X chromosome inactivation or lyonisation. It involves shortening the chosen X chromosome so that it more resembles the Y chromosome, and means that it cannot be fully used by the cell. It's actually the body's way of redressing the balance between males and females, so that both express the same number of chromosomes (rather than females expressing two and males one). Lyonisation happens in every female, but tortoiseshell cats make the process startlingly visible.
It is still possible for male cats to be tortoiseshell, although it's not common. There is a condition, present in humans too, known as Klinefelter's syndrome. This is when a male has three chromosomes, XXY, rather than two. Just like in females, one of the X chromosomes is switched off, which allows for the development of tortoiseshell colouring (but of course, only if the cat has the correct genotype). However, this condition results in males being sterile.
One last point of interest is that all female animals are technically mosaics. Mosaicism is when the cells of an animal contain more than one different genotype. The most common reason for this is X inactivation, and one of the most unusual reasons is that the animal is a chimera (a combination of more than one embryo). There are a number of other causes of mosaicism, and interestingly, brindle dogs are technically mosaics too (see the Brindle page for more on this).
Some other familiar examples of sex-linked gene expression include haemophilia and colour blindness in humans, which affect mainly men.
Haemophilia is a blood clotting disorder caused by a defective recessive gene found only on the X chromosome. Females, having two Xs, follow the usual inheritence rules for this gene. A female with just one copy of it will not express it (she will be a carrier of haemophilia), but a female with two copies will do (she will be a sufferer). However, for males, just one copy of the defective gene will result in haemophilia. This is because in females with one copy of the defective gene and one of the normal gene, enough of the body cells switch off the defective gene (through X chromosome inactivation) to enable the female to display few or no symptoms of the disorder (except in rare cases where the random lyonisation results in more genes switching off the healthy gene, in which case the female may show some symptoms). Men can only have one copy of this particular gene (because it only occurs on X and not Y), so they only have one chance to get it right (although a man with Klinefelter's syndrome, see above, may well be able to escape the problem).
Women may be carriers of haemophilia but men can't be. In order for a man to inherit it, he only needs his mother to be a carrier (and to be a little unlucky - only half of her children will inherit the defective gene, and only half of those are likely to be male. The chance of a woman carrier producing a boy with haemophilia is roughly 1 in 4). For a woman to suffer from haemophilia, she would need a copy of the gene from both parents. This would mean her mother would have to be a carrier and her father would have to be a sufferer, so while female sufferers do occasionally occur, they are very rare.
Weirdly enough, haemophilia actually protects against malaria. In some areas of Africa, the prevalence of malaria means it in a way it can actually be advantageous to be a carrier or sufferer of haemophilia, and as a result the mutation is much more common in these areas than in the general population.
All a bit of a digression, but an interesting one.
Mutation
Mutations occur when there is an error in the DNA replication process. The process is generally very accurate and errors are rare, but they do occur on occasion. The error generally consists of a base being copied incorrectly, for example the codon GTC being copied as GAC. This changes the amino acid that the codon codes for, and hence changes the sequence of amino acids making up the protein, resulting in a slightly different protein (or different structured protein) being produced. This is how different alleles come into existence - through mutations in the normal genes.
The more times a DNA sequence is copied, the more likely it is that a mutation will occur. Therefore, rapidly replicating cells are more likely to produce mutants. Male sex cells (sperm) are replicated many more times than female sex cells (eggs), of which very few are produced in the lifetime of a female mammal. This means that there are many more opportunities for errors in the male. The likelihood of these errors increases with the age of the male.
Natural Selection and Evolution
When a mutation occurs, the new protein may be more or less effective at its job than the original one. Less effective proteins may result in problems for the animal, if the protein is a vital one. For example, sickle cell disease is caused by a mutation in a single base of the gene that codes for haemoglobin, which is an essential part of red blood cells. Red blood cells carry oxygen around the body, and their shape enables them to do this effectively. The single base change that is present in people with sickle cell disease results in a change in the structure of the haemoglobin in the red blood cells, which affects their shape. The new shape is much less efficient at carrying oxygen.
Mutations which result in the animal being less able to survive are not likely to become part of a species' general DNA. The simple reason for this is
that animals with the mutation are likely to die before they have chance to pass on their genes. Animals without the mutation will live longer and reproduce more, so the number of animals of the species without the mutation will eventually outnumber those with it. Sometimes mutations occur which are benefitial to the animal, and these may well become part of the species' DNA, because of the increased survival and reproduction rate of animals with them. The principle is simple
- nature selects for those mutations which are good and removes those which are bad. Sometimes the benefit for the animal may be hard to discern, especially if the mutation results in just a very slight change. However, selection occurs even here, albeit very slowly. A cheetah with slightly longer legs than other cheetahs may be a slightly faster runner. That slight increase in speed will give that cheetah and its descendants a slight advantage when it comes to catching prey. In times
of drought, when prey is hard to come by, these cheetahs have a slightly better chance of survival than other cheetahs. The number of cheetahs with the mutation will slowly increase until it overtakes the number without, and eventually all cheetahs will have it.
As the example of drought shows, changes in the environment can affect which mutations are most useful, and so affect the process of natural selection. Sometimes they simply speed up changes already taking place by making conditions tougher, and other times they can cause completely new changes, such as in the peppered moth (see below), where mutations causing darker wings were selected for due to an environmental change.
The mutations that are selected for aren't necessarily "good" ones, such as in the haemophilia example further up the page. In an area where malaria is common, haemophilia can be the lesser of two evils.
Evolution is the accumulation of genetic changes in a species over time. It is a slow process, as most genetic changes are very minor (like the slight increase in cheetah leg length). Eventually a series of small steps will add up to a big one, and the species will change enough for it to be classified as a new species. Generally, by this point, the old species has disappeared, although sometimes it may continue to exist in isolation elsewhere. Species with a high reproduction rate will evolve faster than those with a slow reproduction rate, for example mice evolve faster than elephants.
Famous examples of evolution include Darwin's finches (diversification - evolution to fill gaps) and the peppered moth (evolution in response to environmental change).
Darwin's finches (so called because Charles Darwin studied them in detail) are a collection of finch species living in the Galapagos. They are all very closely related and originate from just one ancestor. Each species possesses a different size and shape of beak, which enables it to feed on different things. In most other respects, the finch species are very similar. They are an excellent example of how a species has diversified in order to take advantage of different food sources. An individual that is able to feed on something new will do very well for itself, as it will have no competition for its food source. It will evolve first into a variety/sub-species of the original species, and then eventually into a new species.
The peppered moth is generally white with dark patterning, which enables it to blend in with lichen on trees and so makes it less susceptible to predation. However, during the industrial revolution in Britain in the 1800s, pollution caused lichen to die, and the peppered moth was no longer so well camouflaged. A white moth stands out like a sore thumb on a brown tree, so such moths were very likely to be preyed upon. In a small number of individuals, a mutation occured which resulted in a melanic (dark) form. This is something that happens in many species, along with the albino mutation, but generally natural selection goes against the mutation. In the case of the peppered moth, however, these dark individuals became the ones most likely to survive and reproduce. In a very short space of time, the genetic makeup of the species changed so that the majority of peppered moths were dark. As pollution decreased in the 1900s and lichen started to return, this change was reversed and the peppered moth has now mostly reverted back to its white form. Evolution does not often occur this quickly, and when it does it is usually in response to drastic environmental change. In such situations many species become extinct because they are unable to evolve fast enough. They may have a slow reproduction/mutation rate, or they may simply, through bad luck, fail to produce the right mutation in the short period of time they have.

A peppered moth in its usual white phase.
What is a Species?
Officially, if two animals are unable to mate in the wild and produce fertile offspring, they are of different species. However, there are many cases where this doesn't appear to tell the whole story. Some species of bird (particularly ducks and geese) are able to interbreed without problems, as well as many plants, and sterility in many hybrids is a matter of degree, with some simply being less fertile than their parents. Sometimes it even depends on sex - males are more likely to be sterile than females. Perhaps the biggest problem is understanding how exactly species become separate from one another.
The domestic dog has been granted the position of being a subspecies of the grey wolf, and has been given the scientific name canis lupis familiaris (the wolf is canis lupis). A subspecies is a variety of a species that appears to be genetically distinct from the rest, yet is still able to interbreed with others of the species without any problems. In the case of the domestic dog, it seems ludicrous at first to say that a Chihuahua belongs to the same species as the wolf. Apart from the physical difficulties of interbreeding these two members of the species, the Chihuahua bears no visual resemblence to the wolf at all. The red wolf (canis rufus) and the grey wolf seem to have more in common than any wolf does with most domestic dogs, yet these are classified as separate species.
The key in fact lies in the DNA. A great deal of variation may exist in a species' DNA, and this can lead to drastic visual and behavioural differences between subspecies. However, this variation can only be in particular parts of the DNA. Changes such as chromosomes moving to other places in the DNA are enough to create breeding difficulties and so distinguish different species. Most of these species-defining changes are not yet known to science.
It appears that selective breeding by humans, in focusing almost entirely on producing aesthetic and behavioural traits, ignores those changes which would make it impossible for our domestic animals to interbreed with their wild cousins, and which natural selection, by contrast, does tend to favour. A dog's body still works in much the same way as a wolf's body does, even though they appear so different on the outside.
Selective Breeding
Selective breeding is basically intelligent design. It's selection of breeding couples by conscious beings for a purpose, whether that purpose is to create a working dog or a pet guinea pig. Natural selection, by contrast, has no purpose. It simply involves creatures tending to survive more or less well due to their genetic makeup, and so tending to create species and subspecies with similar genetic makeups. There is no conscious selection of particular traits, and as a result, natural selection is much, much slower than selective breeding. A trait can be bred into a species by selective breeding in just a few generations, when it may take hundreds or thousands of years in a wild species.
There is recent evidence that selective breeding for one trait may inadvertantly introduce other traits, so it may not be true that all traits in domestic animals have been specifically bred for.
A long-term study into the domestication of foxes has introduced this surprising idea. Around 35 generations of silver-phase red foxes (vulpes vulpes) have been selectively bred by Russian scientists for just one trait - tameness. The resulting foxes behave very much like dogs, and show some surprising mutations. Most of the mutations are very rare and have only occurred
in a few of the foxes so far, but they include curled tails, white spotting, folded ears and short legs.
The founder of the study, Dmitry K. Belyaev, noticed that most domesticated mammal species show the same sorts of physical changes. Folded ears occur in dogs, cats and pigs, among others, and curly coats are also found in rats, mice, sheep and more. White markings (lack of pigment in particular areas due to partial leucism) appear in most, if not all domesticated mammals. These and other changes seem to be more or less universal, and appear early on in domestication, suggesting that it is something in the process of domestication that causes them. It's unclear exactly what this is, but Belyaev believed
it was due to links between the genes controlling behaviour and the genes controlling development (and so physical appearance). In breeding for domesticity we choose those animals which are most docile, and in doing so we promote changes in the genes controlling behaviour, possibly through selecting for low levels of hormones such as adrenaline. According to Belyaev these changes give rise to a whole series of changes in the genes controlling the animal's development. The study does appear to support this hypothesis.
Partial leucism is a common trait in a number of species of birds that live in close proximity to humans, including ducks and pigeons. Do these white markings occur so regularly just because they're not weeded out by natural selection (living close to humans affords them protection from predators, and provides a stable food source), or are they another example of the link between tameness and colour? We don't currently know, but it's certainly an interesting thought.

Mallard duck with partial leucism. For more on leucism click here

** Please note that I am not a research scientist, and the information on this page comes from my own knowledge and observation of dogs, observational and testing data provided via e-mail by site visitors, any research papers linked on the page, and the information provided by Dr Sheila M. Schmutz on her excellent website http://homepage.usask.ca/~schmutz/dogcolors.html
For further genetics resources, see the Links page